






Conclusion
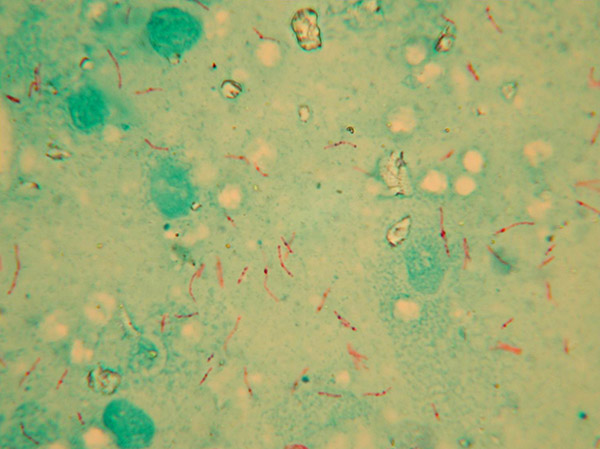
M. bovis is exquisitely adapted for survival in the host for long periods, and establishing chronic infections with a long incubation period that are difficult to detect in the live animal. These same characteristics also allow the organism to survive in the environment for long periods, posing an infection risk to cattle and wildlife from this environmental exposure.
A basic knowledge of the microbiology of mycobacteria is crucial for an understanding of the on-farm risks and the development of disease control strategies.